


纯生信6分+,网络毒理学双污染物×双疾病,思路新颖,换个分子即可复现!
题目:6PPD和6PPD-醌的呼吸毒性机制:基于网络毒理学和分子对接的综合研究
英文名:Respiratory toxicity mechanism of 6PPD and 6PPD-quinone: An integrated study based on network toxicology and molecular docking
杂志:Ecotoxicology and Environmental Safety
影响因子:6.1
发表时间:2025年6月11日
研究背景:6PPD作为广泛使用的橡胶抗氧化剂,主要通过轮胎-路面磨损颗粒释放到环境中,其臭氧衍生物6PPD-Q是城市径流死亡综合征的主要毒性因子。两者通过颗粒物和气溶胶传播,对呼吸系统产生有害影响,但具体致病机制尚不明确。现有研究多聚焦于其神经毒性、生殖毒性和发育毒性,缺乏对呼吸毒性的系统评估,亟需阐明其对人体呼吸系统健康风险的分子机制。
研究思路:本研究整合网络毒理学、分子对接和分子动力学模拟技术,首先通过数据库预测6PPD和6PPD-Q的呼吸毒性潜力,再通过多数据库挖掘筛选两者的潜在靶点,结合GeneCards和OMIM数据库分类的急慢性呼吸损伤相关靶点进行交集分析,构建蛋白质相互作用网络并筛选核心靶点,通过GO、KEGG和DO富集分析解析核心靶点参与的生物学过程和信号通路,借助分子对接验证化合物与核心靶点的结合能力及模式,最后通过分子动力学模拟验证关键靶点复合物的稳定性,从而系统阐明6PPD和6PPD-Q诱导呼吸毒性的分子机制和关键靶点。
研究结果:
1、6PPD诱导急性呼吸系统疾病(ARD)的靶点筛选与富集分析
本研究通过PubChem数据库检索6PPD获取其标准结构(图1A)。最初从BindingDB、ChEMBL、SwissTargetPrediction和TargetNet数据库中筛选出154个6PPD靶点。利用GeneCards和OMIM数据库分别确定肺水肿(1004个)、急性肺炎(892个)、急性肺损伤(ALI,1160个)、急性呼吸窘迫综合征(ARDS,1242个)和急性支气管炎(3139个)的相关靶点,合并去重后共获得4104个ARD相关靶点。通过韦恩图分析鉴定出6PPD与ARD靶点之间共82个潜在交集靶点(图1B),并构建“毒性化合物-靶点-疾病”(TTD)网络图(图1C)。
利用STRING数据库构建包含76个节点和619条边的PPI网络,平均节点度为16.3。使用Cytoscape3.8.2分析PPI网络的拓扑特性(包括度中心性DC、介数中心性BC、紧密中心性CC、最·大团中心性MCC、最·大邻域组件密度DMNC和最·大邻域组件MNC)并可视化网络图(图1D)。根据度中心性,排名前六的靶点为HIF1A、NFKB1、ESR1、SRC、EGFR和PPARG。随后对76个交集靶点进行富集分析。GO分析显示,排名前十的生物学过程(BP)、细胞组分(CC)和分子功能(MF)条目主要涉及信号转导、基因表达调控、代谢过程、蛋白质功能与相互作用以及细胞结构与定位(图1E),这些功能与外源性物质应答和免疫反应密切相关。KEGG分析表明,交集靶点主要参与信号转导和细胞应答相关通路(如MAPK信号通路、PI3K-Akt信号通路、HIF-1信号通路)以及免疫和炎症相关通路(如Th17细胞分化和趋化因子信号通路)(图1F)。所有GO和KEGG富集结果均与6PPD诱导的急性呼吸系统损伤密切相关。接下来,分别从DC、BC、CC组和DC、MCC、DMNC、MNC组中独立筛选核心靶点。具体而言,第一组鉴定出12个重叠靶点(图1G、H),第二组鉴定出1个重叠靶点(图1I)。将两组重叠靶点合并去重后,与GSE68610、GSE261559和GSE283072数据集的差异表达基因(DEG)进行交叉分析,最终鉴定出7个在ARD中显著差异表达的核心靶点:NFKB1、HIF1A、EGFR、RELA、MAPK14、NR3C1和JAK2。筛选结果通过多组环形火山图可视化(图1J)。

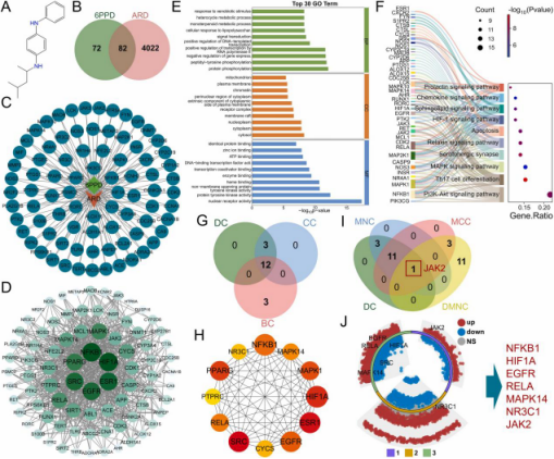
图1
2、6PPD诱导慢性呼吸系统疾病(CRD)的靶点筛选与富集分析
通过GeneCards和OMIM数据库鉴定间质性肺病(ILD,1351个)、慢性肺损伤(CLI,59个)、慢性支气管炎(1143个)和特发性肺纤维化(IPF,1115个)的相关靶点,合并去重后共获得2594个CRD相关靶点。利用韦恩图筛选出6PPD与CRD靶点之间的55个潜在交集靶点(图2A),并绘制TTD网络图(图2B)。通过STRING数据库构建包含50个节点和335条边的PPI网络,平均节点度为13.4。使用Cytoscape3.8.2分析并可视化PPI网络的拓扑特性(图2C)。根据度中心性,排名前六的靶点为HIF1A、NFKB1、EGFR、SRC、PPARG和RELA。随后对55个交集靶点进行富集分析。GO分析显示,交集靶点主要参与信号转导、免疫反应、代谢调控、基因表达调控、氧化应激以及细胞结构与定位(图2D)。KEGG分析表明,交集靶点主要参与免疫反应和炎症(如T细胞受体信号通路、Th1/Th2细胞分化)、信号转导(如HIF-1信号通路、PI3K-Akt信号通路)、细胞代谢与分化(如鞘脂信号通路)(图2E)。这些结果表明,6PPD与CRD的交集靶点主要与免疫反应调控、信号转导、细胞代谢、氧化应激以及神经和生殖系统相关。按照相同流程,分别鉴定出12个(图2F、G)和4个(图2H)重叠靶点。将两组重叠靶点合并去重后,与GSE40839、GSE188464和GSE236857数据集的DEG进行交叉分析,最终鉴定出5个在CRD中显著差异表达的核心靶点:EGFR、PPARG、NR3C1、CYCS和JAK2。筛选结果通过多组环形火山图可视化(图2I)。

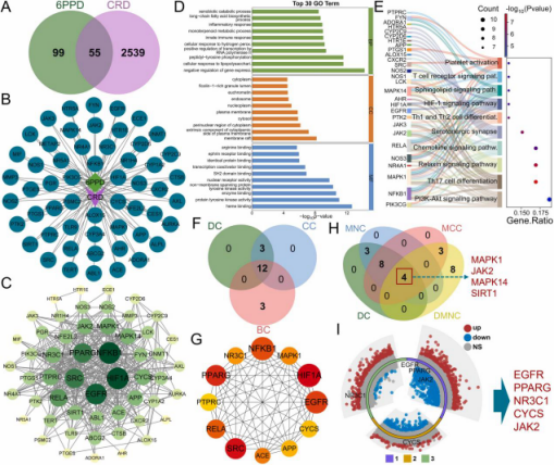
图2
3、 6PPD诱导呼吸毒性核心靶点的分子对接
通过韦恩图分析确定6PPD与ARD和CRD核心靶点的并集(9个)和交集(3个)靶点(图3A)。对这些并集靶点的疾病本体(DO)富集分析显示,核心靶点主要与呼吸系统、消化系统、皮肤、代谢、免疫系统、恶性肿瘤和神经退行性疾病相关(图3B、C),其中呼吸系统疾病主要包括哮喘和支气管疾病。尽管食管炎属于消化系统疾病,但气溶胶颗粒可能吸入并沉积于食管,因此值得关注。对6PPD与3个交集靶点(EGFR、NR3C1、JAK2)进行分子对接研究。AutoDock生成的结果显示,三个靶点的结合能均低于-20.9kJ/mol(-5kcal/mol)。具体而言,6PPD与EGFR的结合能为-26.23kJ/mol(图3D),属于中等结合强度;与NR3C1的结合能为-29.68kJ/mol(图3E),与JAK2的结合能为-30.24kJ/mol(图3F),均显示出强结合能力。这些发现表明6PPD与核心交集蛋白具有显著亲和力,尤其是NR3C1和JAK2,提示它们在6PPD诱导呼吸毒性的分子机制中起关键作用。
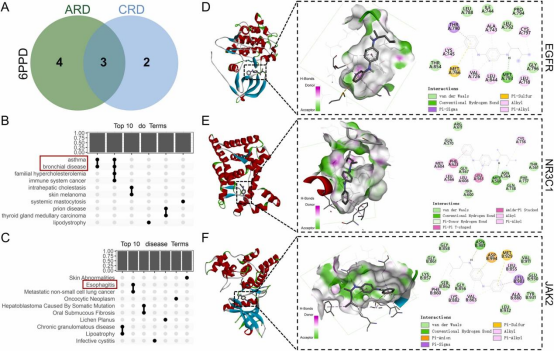

图3
4、6PPD-Q诱导急性呼吸系统疾病(ARD)的靶点筛选与富集分析
本研究通过PubChem数据库检索6PPD-Q获取其标准结构(图4A)。从四个靶点预测数据库中鉴定出200个6PPD-Q的潜在靶点。利用韦恩图筛选并确定6PPD-Q与ARD相关靶点之间的108个潜在重叠靶点(图4B),并基于这些结果构建TTD网络图(图4C)。通过STRING数据库构建包含89个节点和646条边的PPI网络,平均节点度为14.5。使用Cytoscape3.8.2分析PPI网络的六种拓扑特性并可视化网络图(图4D)。根据度中心性,排名前六的靶点为PTGS2、GSK3B、CYCS、RELA、JAK2和ABL1。随后对89个交集靶点进行富集分析。GO分析显示,这些靶点主要与信号转导、细胞增殖、免疫反应、炎症反应、凋亡调控和细胞组分结构功能相关(图4E)。KEGG分析表明,交集靶点主要参与信号转导和细胞增殖(如MAPK信号通路、PI3K-Akt信号通路)、神经功能(如血清素能突触、神经活性配体-受体相互作用)以及免疫反应和炎症(如钙信号通路、凋亡)(图4F)。所有GO和KEGG富集结果表明,6PPD-Q与ARD的交集靶点主要与炎症反应失衡、程序性细胞死亡抑制、细胞功能障碍和神经调控失衡相关。按照上述方法,分别筛选出11个(图4G、H)和1个(图4I)重叠靶点。将两组重叠靶点合并去重后,与GSE68610、GSE261559和GSE283072数据集的DEG进行交叉分析,最终鉴定出9个在ARD中显著差异表达的核心靶点:PTGS2、RELA、MCL1、MMP2、MAPK14、APP、FYN、KDR和CDK2。筛选结果通过多组环形火山图可视化(图4J)。
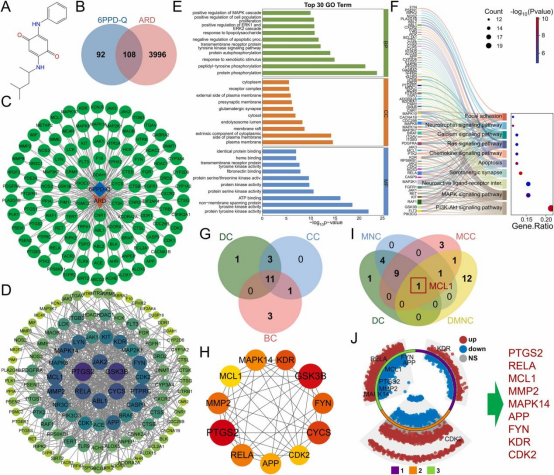
图4
5、6PPD-Q 诱导慢性呼吸系统疾病(CRD)的靶点筛选与富集分析
利用韦恩图鉴定出 6PPD-Q 与 CRD 靶点之间的 83 个潜在交集靶点(图 5·A),并构建 TTD 网络图(图 5B)。通过 STRING 数据库建立包含 71 个节点和 400 条边的 PPI 网络,平均节点度为 11.3。使用 Cytoscape 3.8.2 分析并可视化 PPI 网络的六种拓扑特性(图 5C)。根据度中心性,排名前六的靶点为 PTGS2、JAK2、MMP2、GSK3B、NR3C1 和 RELA。随后对 71 个交集靶基因进行富集分析。GO 分析显示,这些基因主要参与信号转导、免疫反应、炎症反应、神经调控、代谢和细胞功能(图 5D)。KEGG 分析表明,这些交集靶基因主要参与免疫细胞激活与分化(如 T 细胞受体信号通路、Th17 细胞分化)、信号转导(如钙信号通路、Rap1 信号通路)、细胞黏附与迁移(如黏着斑通路)、细胞增殖(如 PI3K-Akt 信号通路)、炎症反应(如趋化因子信号通路)和神经调控(如神经活性配体 - 受体相互作用)(图 5E)。上述富集分析结果表明,6PPD-Q 与 CRD 的交集靶点主要与信号转导异常、细胞黏附与迁移受损、持续性炎症反应以及神经和免疫系统功能障碍相关。按照相同流程,分别鉴定出 11 个(图 5F、G)和 3 个(图 5H)重叠靶点。将两组重叠靶点合并去重后,与 GSE40839、GSE188464 和 GSE236857 数据集的 DEG 进行交叉分析,最终鉴定出 4 个在 CRD 中显著差异表达的核心靶点:JAK2、FYN、NR3C1 和 CYCS。筛选结果通过多组环形火山图可视化(图 5I)。

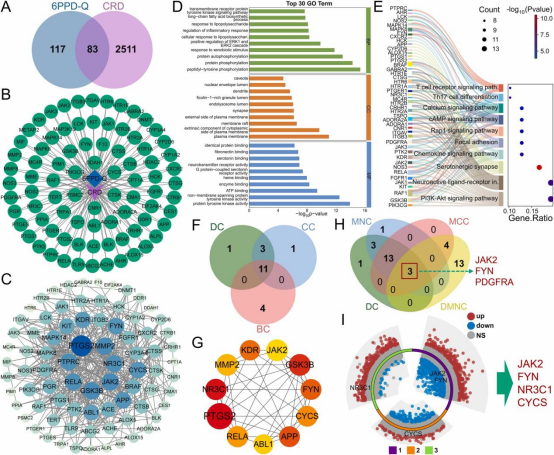
图5
6、6PPD-Q诱导呼吸毒性核心靶点的分子对接
通过韦恩图分析确定6PPD-Q与ARD和CRD相关核心靶点的并集(12个)和交集(1个)(图6A)。对这些并集靶点的DO富集分析显示,核心靶点主要与免疫、神经、血管、呼吸、消化、皮肤和代谢系统疾病相关(图6B、C),其中呼吸系统疾病主要包括鼻窦炎和特发性间质性肺炎。此外,细胞因子风暴作为一种全身性炎症状态,在肺组织中尤为显著,是急性肺损伤相关疾病的主要病因之一(Zhou等,2023)。对6PPD-Q与唯·一的交集蛋白FYN进行分子对接以研究其相互作用(图6D)。AutoDock生成的分子对接结果显示,6PPD-Q与FYN的结合能为-29.01kJ/mol,表明中等结合强度。这种强相互作用提示FYN在6PPD-Q诱导呼吸毒性的分子机制中起重要作用。

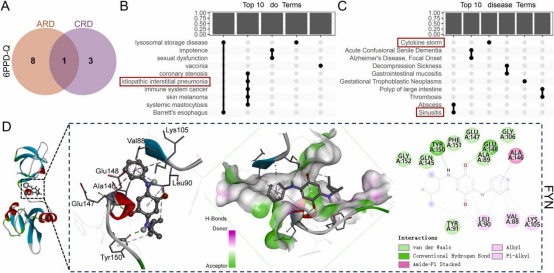
图6
7、6PPD和6PPD-Q核心靶点的联合分析
对6PPD和6PPD-Q诱导急慢性呼吸毒性的核心靶点进行富集分析。GO分析显示,尽管两者均干扰信号转导,但涉及的通路存在显著差异:6PPD优先激活病毒/LPS应答相关通路(如TLR/NF-κB),并通过microRNA双向调控RNA聚合酶II转录;而6PPD-Q特异性靶向VEGFR信号通路,通过FYN介导的肽酪氨酸磷酸化调节膜筏功能。在基因表达水平上,6PPD通过HIF1A/PPARG调控代谢相关microRNA网络,而6PPD-Q通过DNA结合转录因子(如RELA)促进蛋白质核转运,表明两者可能分别通过表观遗传重塑和转录后修饰发挥毒性作用。在免疫反应方面,6PPD诱导的Th17分化和PD-1/PD-L1checkpoint抑制与病毒防御机制相关;而6PPD-Q通过IL-1β生成和IL-17通路激活中性粒细胞浸润,导致急慢性炎症的差异表现(图7A、B)。KEGG通路比较分析进一步证实了这些差异:6PPD主要驱动HIF-1/ROS致癌轴和非酒精性脂肪肝代谢紊乱,其与EGFR的相互作用(-26.23kJ/mol)增强细胞异常增殖;而6PPD-Q显著富集于VEGF/PI3K-Akt通路和动脉粥样硬化网络,其靶点KDR和CDK2协同促进血管渗漏和细胞周期紊乱。值得注意的是,尽管两者均富集于PD-1/Th1-Th2免疫调控模块,但6PPD通过NR3C1抑制糖皮质激素抗炎作用,而6PPD-Q通过APP调控神经退行性通路(图7C、D),这些发现凸显了其毒理学机制的器官特异性倾向。通过韦恩图分析,鉴定出6PPD和6PPD-Q与呼吸毒性相关的5个重叠靶点(呼吸毒性靶点总数:6PPD为9个,6PPD-Q为11个):NR3C1、MAPK14、RELA、CYCS和JAK2(图7E),这些靶点构成呼吸毒性的共同调控枢纽。PPI网络和KEGG富集分析表明,这些靶点通过多层次相互作用驱动共同的损伤机制(图7F、G):具体而言,RELA和JAK2形成转录调控模块,协同激活IL-17/Toll样受体通路,导致Th1/Th17分化失衡和T细胞功能异常;MAPK14-RELA复合物通过TNF/NOD样信号轴触发NF-κB依赖性炎症反应;而CYCS介导的线粒体凋亡与呼吸上皮细胞死亡密切相关。

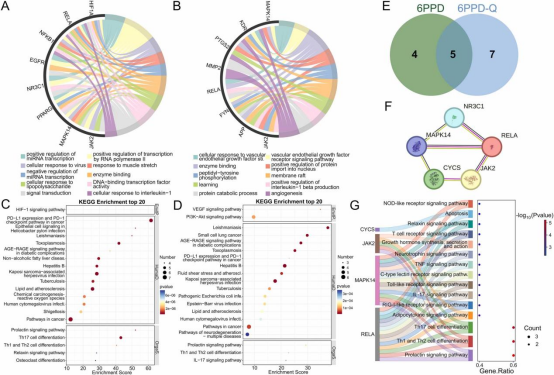
图7
8、呼吸毒性关键靶点的分子对接
由于已完成6PPD与NR3C1和JAK2的分子对接,本研究重点分析6PPD与其他三个关键毒性靶点(RELA、MAPK14、CYCS)的相互作用(图8A-C);而6PPD-Q则与所有五个核心交集蛋白(NR3C1、RELA、MAPK14、CYCS、JAK2)进行分子对接(图8D-H)。AutoDock生成的分子对接结果显示,所有结合能均低于-20.9kJ/mol(-5kcal/mol)。其中,6PPD与CYCS的结合能最·低(-38.37kJ/mol),其次是6PPD-Q与CYCS(-34.94kJ/mol),两者均与CYCS形成两个氢键,提示CYCS可能是毒性化合物诱导呼吸毒性的关键靶点。此外,6PPD与MAPK14(-32.66kJ/mol)以及6PPD-Q与MAPK14(-33.96kJ/mol)的相互作用也显示出强结合强度;6PPD-Q与NR3C1(-28.41kJ/mol)、RELA(-21.93kJ/mol)和JAK2(-27.15kJ/mol)的相互作用为中等结合强度。这些结果(表2)表明核心交集蛋白与6PPD和6PPD-Q均具有强亲和力,尤其是CYCS和MAPK14,提示这些蛋白在两者诱导呼吸毒性的分子机制中起关键作用。

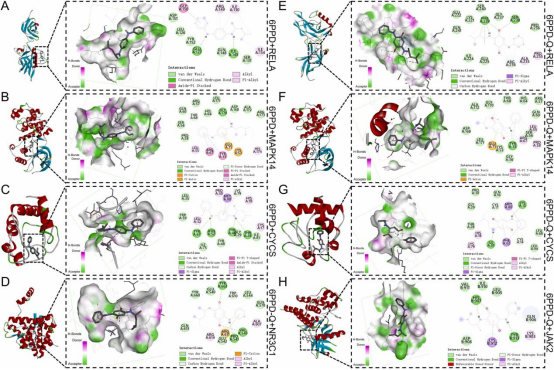
图8
9、关键毒性靶点CYCS的分子动力学模拟验证
基于分子对接实验,CYCS与6PPD和6PPD-Q均显示出显著的结合活性。本研究采用分子动力学模拟(MDS)评估这些复合物的稳定性。CYCS-6PPD复合物的MDS结果(图9A)显示,均方根偏差(RMSD)快速收敛至平衡状态,均方根波动(RMSF)范围为0.05-0.2nm,回转半径(Rg)保持稳定,证实复合物构象的动态稳定性;氢键数量(0-3个)和结合能(ΔE_MMPBSA=-65.08±2.48kJ/mol,表3)进一步验证了CYCS与6PPD之间的强相互作用。相比之下,CYCS-6PPD-Q复合物(图9B)的RMSD波动范围显著增大,RMSF也有所增加(范围为0.05-0.25nm),且Rg值波动明显;此外,氢键数量(0-2个)和结合能(ΔE_MMPBSA=-33.25±3.25kJ/mol,表3)显著低于6PPD复合物。构象重叠分析证实,6PPD可稳定占据CYCS活性口袋内的优势结合位点(图9C);而6PPD-Q分子在模拟过程中表现出多位点分布特征,表明其与CYCS的亲和力不足以维持稳定的结合构象(图9D)。6PPD-Q与CYCS结合呈多位点分布的关键原因在于结构修饰:与6PPD相比,臭氧衍生物6PPD-Q的苯环上新增两个氧原子,形成醌样结构,这种修饰增加了其极性,降低了分子构象灵活性,并增强了吸电子效应(Yuan等,2020),最终导致其与CYCS蛋白疏水性结合口袋的亲和力降低,难以形成稳定结合。

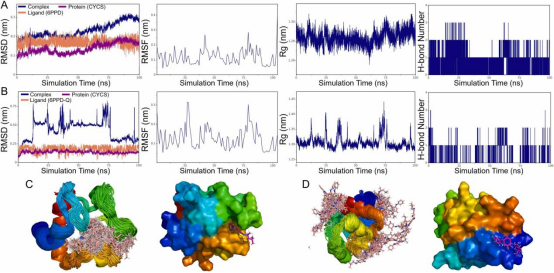
图9
总结:本研究通过多种计算毒理学技术,明确了6PPD和6PPD-Q诱导急慢性呼吸毒性的特异性靶点(6PPD为EGFR,6PPD-Q为FYN)和5个共有靶点(NR3C1、MAPK14、RELA、CYCS、JAK2),证实两者通过破坏线粒体电子传递链、紊乱凋亡通路及激活NF-κB/JAK-STAT炎症级联反应诱导呼吸炎症,其中CYCS和MAPK14为高亲和力关键靶点。研究建立了两者的比较毒理学框架,为环境污染物呼吸毒性的风险评估提供了新方法,也为相关疾病的诊断和防治提供了理论依据。

